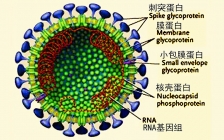
内皮层(皮层内低回声结节)
一、茎尖分区
茎的顶端叫做茎尖,是由叶芽活动形成的。顶芽活动时,生长锥的原生分生组织分裂,向下产生初生分生组织,初生分生组织经初生生长形成初生结构,从而形成茎尖。
(一)分生区
茎尖分生区又称生长锥,一般为半球形,由一团具有分裂能力的原分生组织所构成。
1.原套-原体学说
原套-原体学说将茎尖生长锥分为原套和原体两部分。原套位于其表面,由一层或数层排列整齐的细胞组成。它们进行垂周分裂,扩大其表面积,原体是原套内侧的一团不规则排列的细胞,它们可沿着各种方向进行分裂,增大体积。在营养生长过程中,原套和原体的细胞分裂活动,互相配合,故茎尖顶端始终保持原套、原体结构。大多数的双子叶植物,原套通常是两层,而单子叶植物则有一层或两层。
2.细胞组织分区学说
根据细胞学特征和组织分化动态观察。在裸子植物和已研究的大多数被子植物中发现茎端有分区现象。在原套、原体的中央部位,各有一个原始细胞群,前者称为顶端原始细胞区,后者的称为中央母细胞区。这些细胞较大,并具较大的核和液泡,染色比周围的原套、原体细胞浅,细胞分裂较为频繁。这两群原始细胞分裂形成了围绕在它们周围的周围分生组织区和下方的肋状分生组织区。肋状分生组织区又称髓分生组织区,其细胞较周围分生组织细胞更液泡化,主要进行有规律的分裂,因而形成特殊的“肋状”。有些植物在肋状分生组织区和周围分生组织区上方还有整体如浅盘状的形成层状过渡区,其细胞在茎端纵切面上尉扁平状,排列整齐,如同形成层及其衍生细胞所下形成的形成层带。形成层状过渡区在叶原基形成的间隔期可以有体积的变化。分生区下方形成初步分化的初生分生组织;由原套的表面细胞分化的原表皮层,周围分生组织和肋状分生组织分化形成的基本分生组织和原形成层。
(二)伸长区
伸长区由原表皮、基本分生组织、原形成层三种初生分生组织分化出一些初生组织,其细胞的有丝分裂活动逐渐减弱,伸长区可视为顶端分生组织发展为成熟组织的过渡区域。
(三)成熟区
成熟区的解剖结构特点是细胞的分裂和伸长生长都趋于停止,各种成熟组织的分化基本完成,具备了幼茎的初生结构。在生长季节里,茎尖的顶端分生组织不断进行分裂,伸长生长和分化成熟,结果使节数增加,节间伸长,同时产生新的叶原基和腋芽原基。这种由于顶端分生组织的活动而引起的生长称为顶端生长。
二、双子叶植物茎的解剖结构
(一)双子叶植物节间的初生结构
1.表皮
表皮是幼茎最外的一层细胞,由初生分生组织的原表皮发育而来的初生保护组织。表皮包括表皮细胞、气孔器和各种表皮毛。表皮起着保护和控制蒸腾,防止水分过度散失的作用。有利于幼茎内的绿色组织进行光合作用。
2.皮层
1)外皮层
位于表皮下方,由一至几层厚角组织细胞所组成,协助表皮保护和支持幼茎,其细胞含叶绿体,能进行光合作用。
2)中皮层或皮层薄壁细胞
中皮层或皮层薄壁细胞由体积较大、排列疏松的薄壁细胞所组成,通常含叶绿体,所以,幼茎常为绿色。有些植物幼茎的皮层有分泌道,如棉花、向日葵;后有乳汁管,如甘薯;或有其他分泌结构的分化。有些植物也常含有各种晶体和丹宁等异细胞,如花生、桃等。水生植物茎,一般缺乏机械组织,细胞间隙发达,常有通气组织。
3)内皮层
内皮层通常为幼茎皮层的最内一层细胞,少数植物的内皮层可分化产生凯氏带。有些植物的内皮层富含淀粉粒,称为淀粉鞘,如大豆等。某些木本植物茎的内皮层往往有石细胞群。
3.维管柱
1)维管束
维管束是由原形成层发育而来的束状结构。具有次生生长特性的双子叶植物茎的维管束,包括位于外方的初生韧皮部、位于内方的初生木质部和束内形成层三部分。这种具有束中形成层的外韧维管称为无限外韧维管。一般来说,草本双子叶植物幼茎各维管束之间的距离较大,它们环状排列于皮层的内侧。多数木本植物幼茎内的维管束,彼此间距很小,几乎连成完整的环。
在维管束发育过程中,初生韧皮部由外至内进行向心发育,为外始式。即原生韧皮部靠外方,后生韧皮部靠内方。而初生木质部却由内至外进行离心发育,为内始式。即原生木质部靠内方,后生木质部靠外方。这与根初生木质部的外始式发育顺序,有着根本的不同。在初生韧皮部与初生木质部之间,还具有原形成层保留下老的一层分生组织细胞,称束中形成层,这是进行次生增粗生长的基础。
麻类作物由比较发达的韧皮纤维,其细胞经协同生长和侵入生长而较细长,且其细胞壁是纯纤维素的,故柔软而坚韧,品质最佳,为纺织工业的重要原料之一。
2)髓
髓位于幼茎中央,其细胞体积较大,常含淀粉粒,具有储藏作用。有时,髓细胞也有含晶体和丹宁等的异细胞。
3)髓射线
髓射线位于维管束之间,是连接皮层与髓的薄壁组织,有横向运输和储藏作用。在茎进行次生生长时,与束中形成层相连接的那部分髓射线细胞恢复分裂能力,转变为束间形成层。木本植物茎的髓射线狭窄,草本植物茎的髓射线较宽。
(二)双子叶植物茎节间的次生结构
1.维管形成层的发生、活动与次生维管组织
1)维管形成层的发生
当茎进行次生生长时,首先是位于初生木质部与初生韧皮部之间的束中形成层细胞开始分裂、生长分化,接着与束中形成层相连接的髓射线细胞恢复分裂能力,转变为束间形成层。这样,束中形成层和束间形成层就连成一环,它们共同构成维管形成层。木质化程度高的植物,其维管形成层主要是束内形成层;木质化程度低的草本植物,其维管形成层主要是束间形成层。
2)微管形成层细胞
微管形成层细胞的原始细胞有两种:一是切向面宽、经向面窄的两端尖斜的长梭形细胞,称为纺锤状原始细胞;一是较小、近于等径或稍长的细胞,称为射线原始细胞;前者是形成层的主要成员,沿茎的长抽平行排列,连成一片,发育成茎或纵向(轴向)系统;后者与茎轴垂直排列,分布于纺锤状原始细胞之间,发育成茎的横向(径向)系统。横切面上,纺锤状原始细胞呈扁平长方形,射线原始细胞宽长方形,两者紧密整齐地排列成一环。
3)次生维管组织
维管形成层开始活动主要是纺锤状原始细胞不断进行切向分裂,向外所产生的新细胞径生长,分化形成次生韧皮部,向内所产生的新细胞径生长、分化形成次生木质部(包括导管、管胞、木纤维。木薄壁细胞)。次生木质部和次生韧皮部共同构成次生维管组织,形成茎中纵向的输导系统。同时,射线原始细胞叶不断进行切向分裂,其外侧的新细胞分化形成韧皮射线,内测的新细胞分化成木射线,这两种射线统称为维管射线,成为茎中横向的输导组织,并将次生维管组织隔成许多片区。
4)木材
木材是维管形成层历年产生的次生木质部的总称。
(1)年轮 是指在季节性明显的地区,木本植物的维管形成层在一年中只活动一次而形成的次生木质部。
(2)边材和心材
靠近形成层部分的木材是近几年形成的次生木质部,颜色浅,具有木薄壁组织,其导管,管胞和木薄壁组织有效地担负输导和储藏的功能,这样的木材称为边材。边材木质疏松,木质差。靠近茎中央部分的木材是形成较久的次生木质部,这部分颜色较深,其导管和管胞由于侵填体形成,并因丹宁、树脂、色素等有机物的积累而失去输导功能,这样的木材称为心材。心材木质硬、木质好。
2.木栓形成层的发生、活动与周皮
1)木栓形成层发生
木栓形成层最初发生的位置因植物种类不同而异。有的起源于表皮(苹果、梨等),有的起源于近表皮的皮层薄壁组织(马铃薯、桃等)或厚角组织(花生、大豆等),有的起源于皮层较深处的薄壁组织,有的甚至在初生韧皮部中发生。大多数植物茎中,木栓形成层的活动期是有限的,通常生存几个月就失去活力,再形成新的周皮。这样,木栓形成层的位置则渐向内移。
2)木栓形成层的活动
木栓形成层主要进行切向分裂,向外产生的新细胞经生长,分化为木栓层,向内产生的新细胞经生长,分化为栓内层。
3)皮孔
皮孔是枝条外表上产生的一些呈圆形,椭圆形或枝状的褐色小突起,是茎与外界进行气体交换的通道,也是病虫入侵的“门户”。皮孔一般产生于原来气孔的位置,气孔处或其内方的木栓形成层不形成木栓细胞,而形成许多球形的薄壁细胞,其细胞排列疏松,细胞间隙发达,称为补充组织。随着补充组织越来越多,向外突出,形成裂口便有了皮孔。
树皮 通常意义上的树皮是维管形成层意外所有组织的总称,除了次生韧皮部外,还包括历年产生的周皮和一些已死的皮层、韧皮部等,是茎内同化产物的输导途径,又具有极强的保护功能。
3.双子叶植物茎的次生结构
周皮 位于茎的最外方,由木栓层、木栓形成层和栓内层构成。周皮上通常有皮孔,是老茎进行气体交换的通道。
皮层 位于周皮的内方,含有少量的薄壁细胞,棉皮层中有分泌腔分布,有些植物有异细胞。
初生韧皮纤维 位于皮层内方,呈束状分布。初生韧皮部由于受到内部挤压,除发达的初生韧皮纤维外,其余全被挤毁。
次生韧皮部 位于初生韧皮纤维内方,常与呈三角形或喇叭形的韧皮射线相间隔,由韧皮薄壁细胞、筛管、伴胞、韧皮纤维组织。其韧皮纤维发达;具有输送有机养分和机械支持作用,是次生维管组织的重要组成部分。
维管形成层 位于次生韧皮部内方,由纺锤状原始细胞和射线原始细胞组成。在横切面上,细胞呈扁平长方形或正方形。
次生木质部 位于形成层内方,由导管、管胞、木薄壁细胞、木纤维组成,木纤维发达。起输水分、矿物质和机械支持作用,是次生维管组织的重要组成部分。

初生木质部 位于次生木质部内方,为内始式;是初生结构保留下来的相对完整的结构部分之一。
髓 位于茎的中央,由薄壁细胞组成;常含有淀粉粒等储藏物质;有的髓边缘常有环状的环髓带。
维管射线 包括木射线和韧皮射线。位于次生木质部中的射线称为木射线,位于次生韧皮部中的射线称为韧皮射线,均由薄壁细胞组成,呈径向排列,是横向的输导和储藏组织。
三、单子叶植物茎的解剖结构
(一)禾本科植物茎的解剖结构
1.表皮
表皮是位于茎最外面的一层细胞,由长细胞,短细胞和气孔器有规律地排列而成,外覆角质膜。长细胞是构成表皮的主要成分,其细胞壁厚,不仅角化且硅化,其纵向壁常呈细密的波浪状。短细胞有两种类型,即栓细胞和硅细胞。它们位于两个肠细胞间,排成整齐的纵列。栓细胞的细胞壁栓化,硅细胞含有大量的二氧化硅,硅酸盐沉积于细胞壁上的多少,与茎秆强度和对病虫害抵抗力的强弱有关。
2.基本组织
表皮以内,维管束之间的所有区域皆为基本组织,主要是由薄壁细胞组成。
3.维管束
维管束为有限外韧型,散生于基本组织中。其排列方式有两类:一类以水稻和小麦等为代表,各维管束大体上排列为内、外两环。外环维管束较小,位于茎的边缘内,其大部分埋藏于机械组织中;内环维管束较大,为基本组织所包围。水湿生植物茎的两环维管束之间有大的通气道,形成良好的通气组织,节间中部常形成髓腔;一类是玉米、甘蔗和高粱等,其茎内为基本组织所充满,维管束分散排列于其中;近边缘的维管束较小,相距较近,靠中央的维管束较大,相距较远。维管束的结构组成包括维管束鞘、初生韧皮部。初生木质部位于维管束的近轴方,整个横切面的轮廓呈V字形。V字形的基部为原生木质部,包括1至几个环纹或螺纹导管及少量木薄壁细胞。在分化成熟过程中,这些导管常被拉短、挤破,其四周的薄壁细胞互相分离形成气隙。在V字形的两臂上,各有1个后生的大型孔纹导管。在这两个导管之间充满薄壁细胞,有时也有小型的管胞。初生韧皮部位于初生木质部外侧,其中原生韧皮部已被挤毁,后生韧皮部由筛管和伴胞组成,筛管大而多边形。每个筛管旁边有三角形或长方形的小细胞,叫伴胞。
(二)禾本科植物茎的初生增粗生长与居间生长
1.初生增储生长
禾本科植物的维管束没有束中形成层,不能进行次生增粗生长。但是,甘蔗和高粱等茎实际上比具有次生生长的棉花、花生等的茎还粗壮,其原因是在玉米和甘蔗等茎尖的正中纵切面上,可以看到在原叶基的下面有多由顶端分生组织衍生而来的、扁平的、初生分生组织细胞,它们有规律地排列为垂周行列,这称为初生增粗分生组织。它们快速进行平周分裂,衍生出大量薄壁组织细胞,形成粗壮的牙尖,随着细胞的分裂和增大,形成粗壮的茎。
2.居间生长
禾本科植物每个节间基部都保持幼嫩的生长环即居间分生组织,它们的细胞进行分裂、生长和分化就使每个节间伸长,这称为居间生长。拔节是居间生长的表现,抽穗是几个节间同时进行居间生长的结果。
(三)非禾本科植物茎的解剖结构
表皮 多数单子叶植物茎的表皮终身有效,没有周皮形成。
基本组织 单子叶植物茎表皮以内通常为数层厚壁组织,厚壁组织内侧为基本组织,基本组织内分布着维管束。
维管束 单子叶植物维管束均为有限维管束,维管束中没有形成层,不能进行次生生长,其结构与禾本科植物类似。
四、裸子植物茎的解剖结构
(一)松柏类植物茎的初生结构
表皮 裸子植物的表皮是位于茎最外一层细胞,为初生保护组织,由原表皮发育而来。
皮层 位于表皮与维管柱之间,由基本分生组织分化而来,由多层薄壁细胞组成。皮层内常分布着树脂道。
维管组织(柱) 松柏类植物茎的韧皮部没有筛管、伴胞和纤维,而由筛胞和韧皮薄壁细胞构成。木质部中没有导管、纤维,仅有管胞和木薄壁细胞组成。
(二)松柏类植物茎的次生结构
松柏类植物茎的次生结构,有发达的次生构造,形成木材和树皮。指示组成木质部与韧皮部的成分不同,因此,木材的构造也有差异。
松柏类植物茎次生木质部结构均匀,构造简单,一般含有大量管胞、少量慕薄壁组织、木射线与树脂道等,木质部没有导管和纤维;在晚材中只含有管胞状纤维。有的种类的次生木质部完全由管胞和及少量薄壁组织组成。有的种类木质部、韧皮部、皮层等部位具有树脂道,纵横排列连接成一个系统。由于次生生长形成的木材主要由管胞组成,因而木材结构均匀细致,易与双子叶植物的木材区分。木材中亦有年轮、心材和边材的分化。在木材的横切面上,可看到呈辐射排列的单列细胞的木射线,只有少数种类含有两列细胞的木射线。而双子叶植物茎的次生木质部中通常是单列木射线和多列木射线同时存在。
松柏类植物茎的次生韧皮部中含有筛胞、韧皮薄壁细胞和韧皮射线,也有些种类含有韧皮纤维,有些种类还含有围绕着树脂道的薄壁组织。松柏类植物茎的次生韧皮部中没有筛管和伴胞。
木麻黄等草本裸子植物茎的初生结构包括表皮、皮层和维管柱三部分。表皮层细胞外壁厚,皮层外层细胞局部发育成厚壁机械组织,皮层薄壁细胞中含叶绿体,外韧维管束,具有束内形成层,初生韧皮部由筛胞和位于外围的厚壁组织所组成,初生木质部主要由管胞分子所组成。一般具有次生生长和次生结构。



文章评论(0)